Size limits the sensitivity of kinetic schemes
이거 왜 읽었나?
Journal reading - Stochastic thermodynamics for biological functions
위 리뷰 논문의 레퍼런스라서 읽었다.
세포의 sensitivity가 Markov chain의 topology와 관련있다는 내용이라고 하길래, 흥미로워 보였다.
Overview
Continuous time Markov chain (Markov jump process, kinetic scheme)에서 어떤 관측량의 감도(Hill coefficient)가 perturbation의 support의 size m에 의해 bound된다는 것을 증명한 논문.
평형계에서
또한 이 bound를 포화시키는 새로운 비평형 메커니즘인 nested hysteresis를 제안한다.
Link to PDF and DOI
- DOI: https://doi.org/10.1038/s41467-023-36705-8
- PDF: Size limits the sensitivity of kinetic schemes.pdf
용어 정리부터
FIG1.
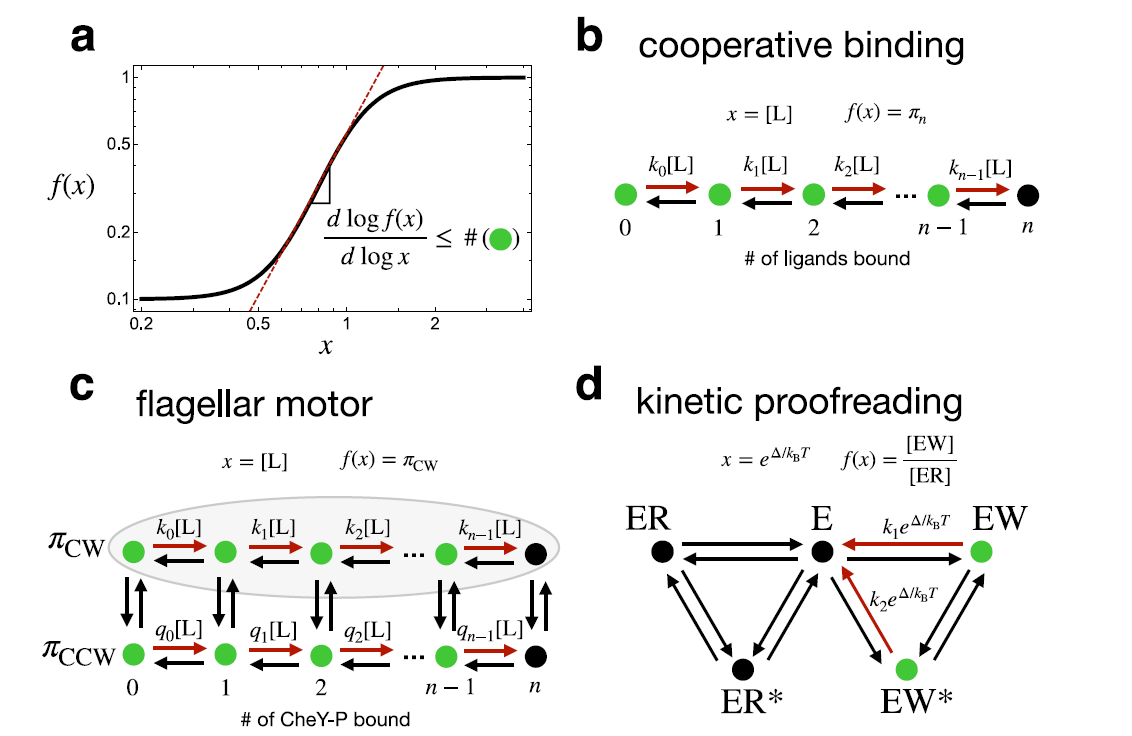
- 기본 모델로 continuous time Markov Process 를 이 논문에서 사용한다.
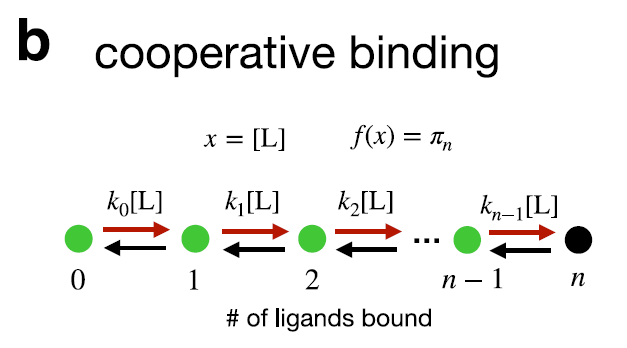
- 이 논문에서 다루는 Markov state이란,
ligand가 붙을 수 있는 효소나 DNA의 상태를 의미한다.
이것에 ligand가 붙은 양상, 효소의 어느 자리에 무슨 ligand가 붙었는지가 달라지면 상태가 달라지는 것이다.
예를 들어 구분 불가능한 ligan binding site가 n개 있는 효소의 상태는, 위의 그림 처럼 ligand가 붙은 수에 따라 0부터 n 까지의 상태로 구별된다.
- 이 논문에서 다루는 perturbation 이란,
주로 ligand concentration을 의미한다. Ligand가 몇 개나 효소에 붙었는지에 따라 효소의 상태가 달라지고, ligand가 붙는 속도는 ligand concentration에 선형적으로 증가한다. 때문에 Markov chain의 transition rate는 ligand concentration에 따라 달라진다. 그래서 위 그림에 state transition rate가
로 표기된 것이다.
결과적으로 평형상태에서 특정 상태에 있을 확률은 ligand concentration이라는 perturebation에 의해 달라진다.
*다른 자료에서 따온 이미지*
다른 자료에서 따온 이미지
- Hill coefficient와 cooperative binding→ 이 논문의 출발점.
일부 효소는 여러 개의 ligand binding site를 가지고 있다. ligand concentration이 증가할 때, 효소의 모든 ligand binding site가 차지되어 있을 확률,
는 시그모이드함수로 증가한다.
이 시그모이드 관계를 정량화하기 위해 Hill function을 사용한다:
여기서
는 ligand concentration, 는 effective dissociation constant, 는 Hill coefficient로 반응의 logarithmic sensitivity를 정량화한다. 가 클수록 입출력 관계가 스위치처럼 날카로워진다. 열역학적 평형 상태에서 는 동시에 결합 가능한 ligand의 최대 수 을 초과할 수 없다는 것이 알려져 있다.
Hill function을
- Support bound → 이 논문에서 제시하는 가장 핵심 개념
perturbation x의 support는 Markov chain을 이루는 state의 부분집합을 의미하는데, “perturebation이 없을 때보다 있을 때 탈출 속도가 더 빨라지는 states”로 이루어진 집합이다. 더 명확히 하면 “x가 증가함에 따라 exit rate가 증가하는 states”이다.
이 논문의 제목에서 있는 size는 곧 support의 개수를 의미한다.
Support bound는 perturbation x의 support size가 logarithmic sensitivity를 bound한다는 개념이며, 이 논문의 핵심 발견이다.
임의의 관측량 A와 B가 있다고 치자. Steady state에서 관측량의 평균을
라고 나타내겠다. m이 perturbation x의 support size일 때 다음 부등식을 만족한다는 것이 논문의 주장이다.
m은 오로지 markov chain의 구조에 의해 정해지기 때문에, A와 B를 어떤 관측량으로 정하는 지와는 무관하다.
앞에서 Hill function을 다룰 때는 ‘효소의 모든 ligand binding site가 차지되어 있을 확률’
식 (7)에서 만약 관측량
Support Bound 증명: 식 (28)~(32)
식 (7)이 왜 성립하는지 증명 흐름을 따라가 보자. 핵심 도구는 Markov Chain Tree Theorem이다.
식 (28): Steady-state를 spanning tree로 표현
Steady-state 확률
식 (29): ⟨A⟩/⟨B⟩는 유리함수다
분자의 최고·최저 차수를
식 (30): 로그 감도를 미분으로 표현
식 (29)를 로그 미분하면:
괄호 안의 각 항은 다항식의 가중 평균 차수이므로, 첫째 항은
식 (31): 차수 범위로 bound
위 범위를 이용하면:
식 (32): Support가 차수를 제한한다 → Support Bound 완성
식 (28)에서 spanning tree는 각 vertex마다 나가는 edge가 최대 하나 (root 제외). 따라서 한 monomial에서
이것이 바로 식 (7)의 support bound다. 증명 어디에도 A와 B의 구체적인 값이 개입하지 않으며, 오로지 그래프의 구조만이 bound를 결정한다.
그래서 이 논문의 목표는
평형계에서 logarithmic sensitivity 부등식을 비평형계로 확장하는 것이다.
주의) 정상 상태와 평형 상태는 동일한 개념이 아니다. 비평형 정상 상태가 있기 때문.
이 논문은 비평형 정상 상태를 다룬다. Steady-State vs Equilibrium State
- 평형계에서는 Hill coefficient ≤ n (Ligand 결합 자리 수)
- 비평형계에서는 Hill coefficient ≤ m (support 크기)
이를 더 구체적으로 이해하기 위해 식 (7)의 corollary인 식 (8)을 보자. observable A를 상태 집합
우변의 구조가 식 (6)의 Hill function 감도
즉, support size m이 Hill coefficient의 역할을 한다.
한편 평형계에서는 식 (10)이 성립한다:
여기서
핵심은 m은 n보다 클 수 있다는 것이다.
따라서, 비평형계에서는 평형계에서 기대되는 것 보다 더 큰 sensitivity가 나타날 수 있다.
는 것이 논문의 주장이다. 이것이 맞음을 실험 데이터를 근거로 설명한다.
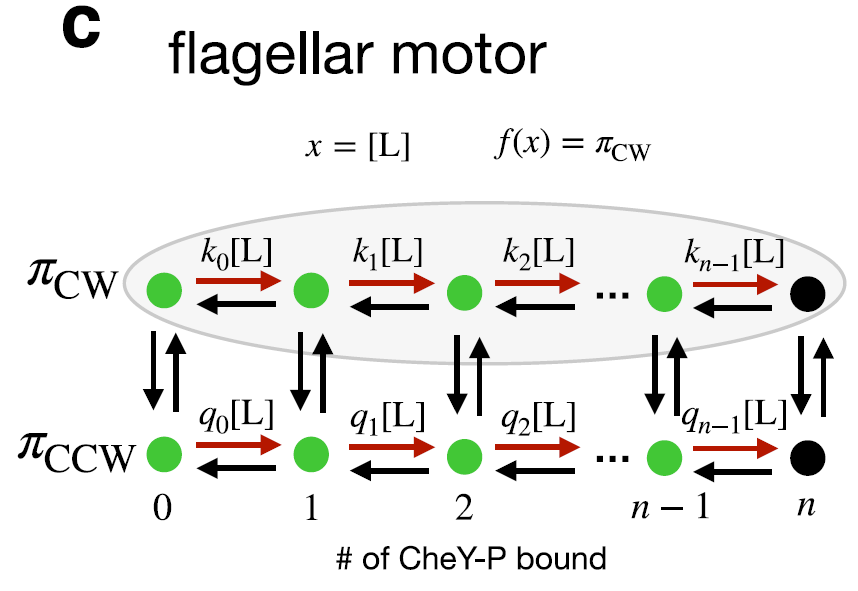
- 응용 사례: E. coli 편모 모터 스위치(비평형 MWC 모델), kinetic proofreading, 화학 농도 sensing
Nested hysteresis → support bound를 포화시키는 비평형 메커니즘
FIG3
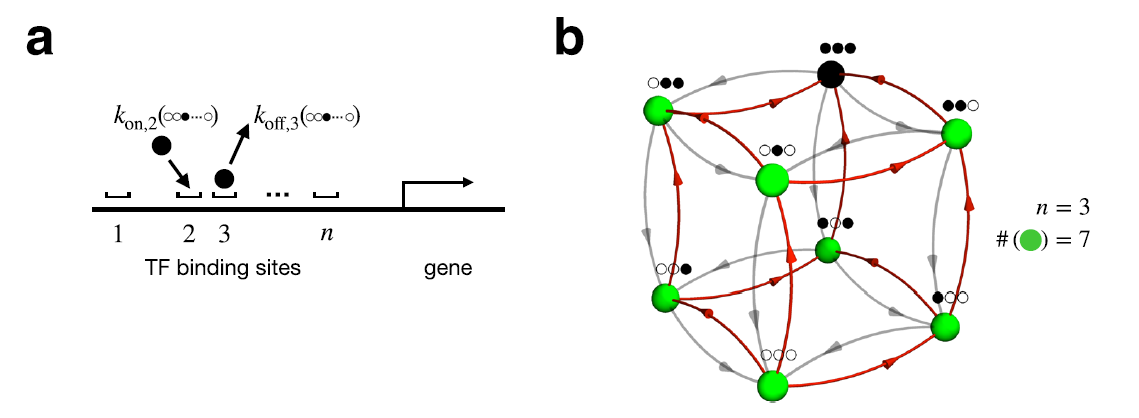
DNA에 여러개의 transcription factor(TF)들이 붙는 상황을 가정하라.
이때 TF는 한 가지이지만, n개의 binding sites는 구별된다.
이런 시스템을 unordered binding 모델이라 부르자.
즉
이 경우 n개의 site 각각이 ligand on/off 두 가지 상태를 가지므로, 가능한 상태의 수는
Fully bound state는 exit rate가 ligand concentration과 독립적이다.
(오직 dissociation constant에 의해서만 exit rate가 정해진다.)
하지만 나머지 모든 상태들은 적어도 하나의 exit edge는 ligand concentration에 dependent하다. 따라서 support size는
Bound 자체는 이렇게 정해졌는데, 이렇게 ligand bound site 개수에 exponential하게 sensitivity가 증가하는 게 가능한가? 그러니까,
이 논문은 위와 같은 경향을 따르는 nested hysteresis라는 간단한 메커니즘을 고안했다.
그리고 이 메커니즘이 실제 DNA와 TF biding을 잘 모사할 수 있는 생물학적 근거 또한 갖추고 있다고 자부한다.
Nested hysteresis는 hyper cybic에 추가로 아래 두 특징을 가진다.
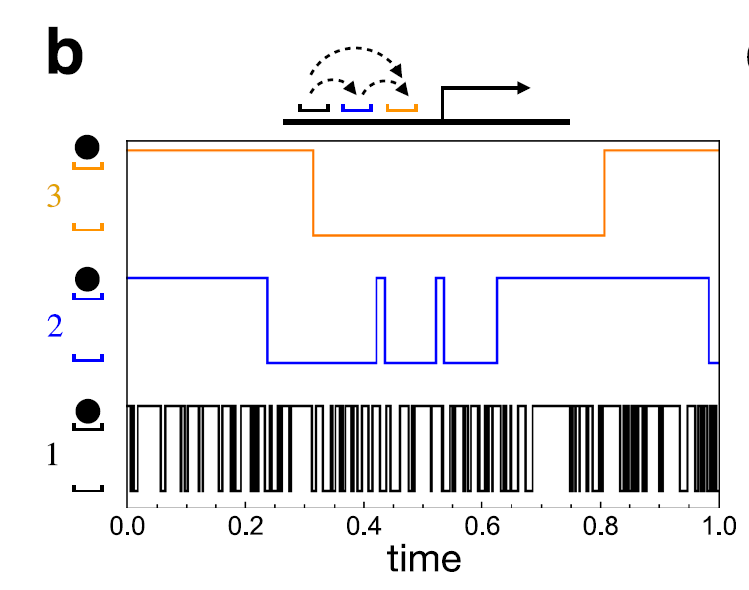
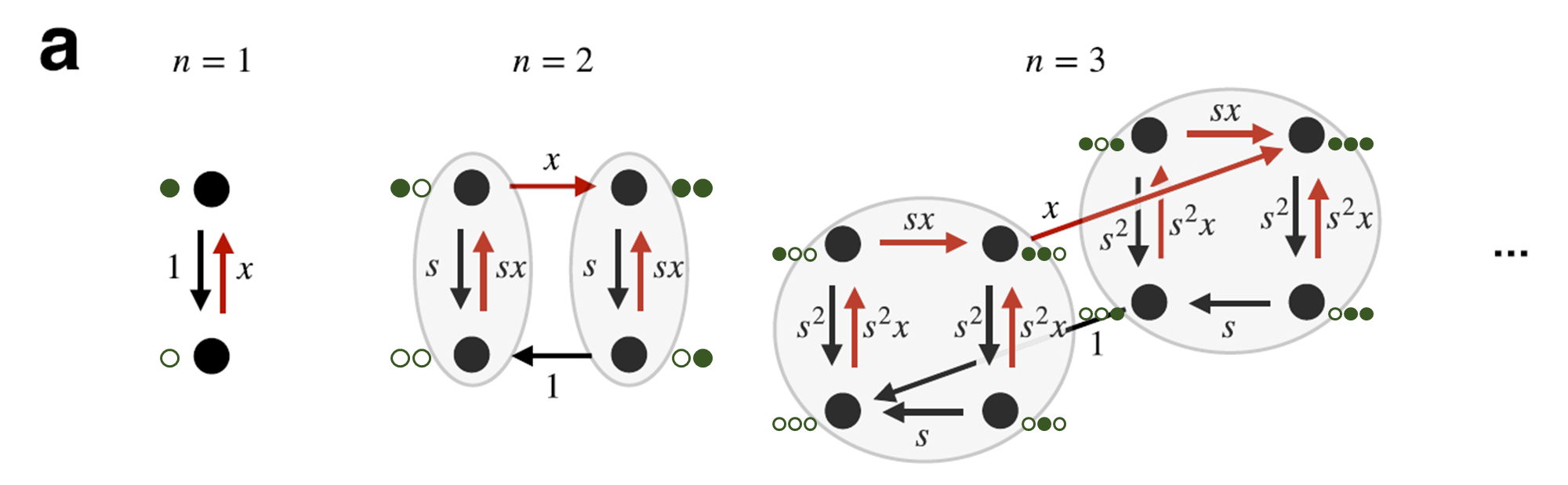
① 결합 자리에 따른 시간 척도의 위계(hierarchy of timescales)
결합 자리에 번호를 붙이면, 높은 번호의 자리일수록 결합/해리가 훨씬 느리게 일어난다.
느린 정도가 꽤나 커서, 다른 결합 자리는 아예 다른 time scale에서 사는 정도이다.
② 중첩된 조건부 결합 규칙
자리
이 두 조건이 결합되면 강한 시간척도 분리 극한에서 각 자리의 steady-state 점유 확률이 다음과 같이 유도된다:
즉 자리마다 유효 Hill coefficient가
여기에 두 극단 상태(fully bound, totally empty)에서 나가는 전이율을
이 식은
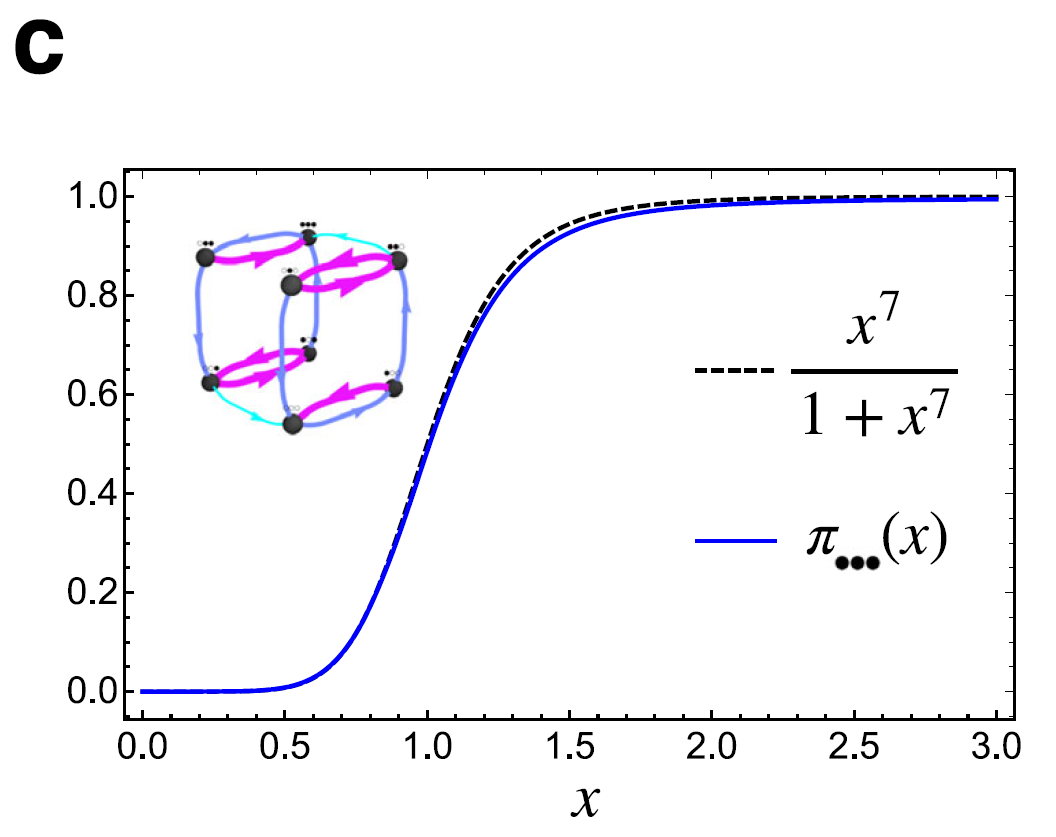
이런 nested hysteresis를 직접 시뮬래이션한 결과, Hill coefficient = 7 의 이론 함수와 잘 맞아 떨어지는 그래프를 얻었다.
생물학적 함의: 평형계에서
실제 실험 데이터를 보면 이 이론에 힘이 실린다.
Hunchback 이라는 유전자의 TF에 대한 sensitivity를 측정한 결과, Hill coefficient가 5-7 정도의 수치로 나왔다. 그런데 TF binding site를 유전적으로 없애버리는 실험을 한 결과, 5개보다 적은 binding site가 있는 것으로 나타났다. 따라서 평형 상태의 모델로는 실험에서 보인 강력한 Hill coefficient를 설명할 수 없다. 대신 binding site에 대해 지수적으로 Hill coefficient가 증가하는 nested hysteresis는 이를 설명할 수 있다.
Questions & Insights
-
Q: 논문이 f(x) 대신 ⟨A⟩π/⟨B⟩π에 집중하는 이유는?
A: f(x)는 너무 막연해서 bound 증명이 불가능. ⟨A⟩/⟨B⟩ 형태로 쓰면 Markov chain tree theorem에 의해 유리함수임이 보장되어 분자·분모의 최고차수 비교로 감도의 상한을 유도할 수 있음. 동시에 이 형태가 실험적 측정량 거의 전부를 포괄해 일반성도 잃지 않음. -
Q: A와 B가 임의적으로 정해질 수 있는데 부등식이 의미 있는가?
A: m이 A와 B로부터 완전히 독립적으로 결정되기 때문에 가능. m은 시스템의 그래프 구조(어떤 상태가 x에 의존하는 탈출률을 갖는가)에서만 나오는 양. Markov chain tree theorem에서 spanning tree는 각 vertex마다 나가는 edge가 최대 하나이므로, x에 의존하는 전이율이 한 monomial에 들어갈 수 있는 횟수가 support 크기 m을 넘지 못함 → πk의 x에 대한 최고 차수가 m 이하 → 어떤 관측량을 고르든 감도가 m을 초과할 수 없음. A, B가 임의적이라는 점이 오히려 이 bound를 모든 측정량에 적용되는 보편적 구조적 한계로 만듦.
-
Q: 편모 모터 MWC 모델에서 CW↔CCW 전환에 ligand가 직접 관여하지 않는데 어떻게 support에 포함되는가?
A: Support는 “어떤 전환이 x에 의존하는가”가 아니라 “어떤 상태의 탈출률(exit rate)이 x에 의존하는가”로 정의됨. CW 상태들은 CW↔CCW 전환(검은 화살표, x 무관) 외에도 CheY-P 결합(빨간 화살표, x 의존) 경로가 있어서 전체 탈출률 = k_i+ (무관한 것들)이 되고, 이 오르면 탈출률이 바뀜 → support 포함. 유일하게 빠지는 상태는 fully bound state(결합 수 = n)로, 더 이상 결합이 일어나지 않아 x 의존 전이가 없기 때문. 따라서 m = 2n.
Related Concepts
Steady-State vs Equilibrium State
더 읽어보고 싶은 레퍼런스
Journal reading - A Flow-Graph Formula for the Stationary Distribution of a Markov Chain
→ Markov chain tree theorem이 뭔지 알 수 있다고 한다.